導入
解糖( γλῠκὖς glykýs 「甘い」およびλύσις lýsis 「溶解」)またはエンブデン・マイヤーホフ・パルナス経路は、グルコースの同化とエネルギー生成のための代謝経路です。それは細胞の細胞質(またはサイトゾル)で起こります。その名前が示すように、グルコースを必要とし、ピルビン酸を生成します。後者は、好気環境では真核生物のミトコンドリアまたは細菌の細胞質で起こるクレブス回路に入るか、嫌気環境では発酵によって代謝されて乳酸塩やエタノールなどを生成します。
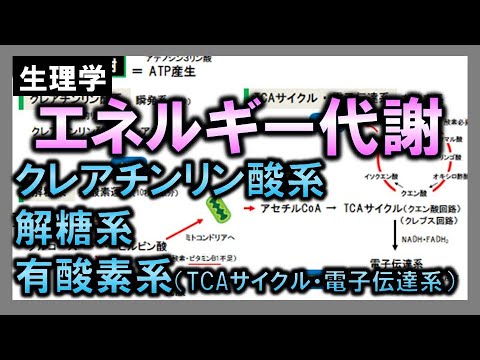
一般原則
解糖は酸素を必要としない ATP 再生メカニズムです。このプロセス中に、私たちは次のことを目撃します。
- 電子受容体 (補酵素 NAD) が還元される酸化還元反応:
- NAD + + 2 H + + 2 e − → (NADH,H + )
- ATP は、ADP のリン酸化によって合成されます (4 つの ATP 分子が形成されますが、2 つ消費され、合計2 つの ATP 分子が形成されます): 2 ADP + 2 Pi + 2 H + → 2 ATP + 2 H 2 O。記号 Pここで、 iは無機リン酸塩HPO 4 2- (またはリン酸水素塩)を表す。
解糖は補酵素の還元を引き起こすため、有機分子の酸化を伴います。これはグルコースのピルビン酸への酸化に相当すると言えます。
- C 6 H 12 O 6 + 2 NAD + → 2 CH 3 -CO-COOH + 2 (NADH,H + )
と組み合わせて
- 2 ADP + 2 Pi + 2 H + → 2 ATP + 2 H 2 O
または合計で
- グルコース + 2 ADP + 2 Pi + 2 H + + 2 NAD + → 2 ピルビン酸* + 2 ATP + 2 (NADH,H + ) + 2 H 2 O
* 「ピルビン酸」(CH 3 -CO-COO – ) という用語は、厳密にはその共役酸(ピルビン酸CH 3 -CO-COOH) を指します。
グルコース-6-リン酸の異性化
これは異性化であり、グルコース-6-リン酸からフルクトース-6-リン酸を生成するホスホヘキソースイソメラーゼによって触媒される可逆反応です。
フルクトース-1,6-二リン酸の合成
この反応はホスホフルクトキナーゼ (PFK) によって触媒され、不可逆的であり、Mg 2+に依存します。この酵素は解糖の最初の特定のステップを触媒します。これは、解糖の「有用な」最終生成物である遊離ATP(遊離ATPはマグネシウムと複合体を形成していないATPの形態)によってアロステリックに非常に強く制御されています。遊離ATP の濃度が高いほど、この反応は遅くなり、逆に、遊離ATP の濃度が低いほど、酵素の活性は高くなります。これは解糖を自己制御するためのサイバネティックシステムです。解糖のいくつかの数学的モデルが開発されており、このステップが解糖の流れを制御するステップの中で最も重要であることが示されています。

ATP による阻害は AMP によって可逆的であるため、ATP/AMP 比を一定に保つことが可能になります。
しかし、それは主にフルクトース-2,6-二リン酸(F26BP) によって調節されています。実際、F6P からの F2,6BP の生成には、F6P 経路の飽和 (「完全すぎる」) を強調するという唯一の機能があります。代謝的な運命を持っています。したがって、アロステリーにより、F26BP はホスホフルクトキナーゼを活性化して F6P の消費を刺激し、それによって自身の形成を防ぎます。
トリオースリン酸の形成
グリセルアルデヒド-3-リン酸(3-PGA)とジヒドロキシアセトンリン酸(PDHA)の形成
この反応は可逆的であり、アルドラーゼ (リアーゼ グループ) によって触媒されます。 (ジヒドロキシアセトンリン酸は一番下の分子です)。トリオースリン酸イソメラーゼのおかげで、D-グリセルアルデヒド-3-リン酸(3-PGA)からジヒドロキシアセトンリン酸(PDHA)に可逆的に切り替えることが可能です。これはアルドール縮合とは逆の反応です。

トリオースリン酸の異性化
この反応は可逆的です(トリオースリン酸イソメラーゼによって触媒されます)が、次の反応では D-グリセルアルデヒド-3-リン酸が消費され、平衡は後者の合成の方向にシフトします。 (以下の画像では、丸で囲まれた P 記号は -PO 3 2-グループを表します)。
エネルギー回収
1,3-ジホスホグリセリン酸の合成
この酸化還元反応は可逆的であり、D-グリセルアルデヒド-3-リン酸デヒドロゲナーゼ(オキシドレダクターゼ)によって触媒され、高い転移能を有するアシルチオエステル結合の形成をもたらします。このステップは解糖の第 2部分の開始部分を構成します。高い移動電位を持つ結合に含まれるエネルギーは、ATP の合成に使用されます。補酵素は還元されます(電子の獲得)。
赤血球では、ムターゼ触媒反応により、ヘモグロビンの重要なアロステリックエフェクター(酸素に対する親和性を調節する)である 1,3-ジホスホグリセリン酸から 2,3-ジホスホグリセリン酸が生成されます。次に、2,3-ジホスホグリセリン酸は、解糖経路に従う 2,3-ジホスホグリセリン酸ホスファターゼによって ATP 分子の生成 (無機リン酸の放出)を伴わずに3-ホスホグリセリン酸に変換されます。
3-ホスホグリセリン酸の合成とATPの回収
ATP の合成 (エネルギー回復) があり、この反応は可逆的で、ホスホグリセリン酸キナーゼ (トランスフェラーゼ) によって触媒されます。
2-ホスホグリセリン酸の合成
この可逆反応は、ホスホグリセリン酸ムターゼ (トランスフェラーゼのグループ) によって触媒されます。
ホスホエノールピルビン酸の合成
この反応は、可逆的エノラーゼ(リアーゼのグループ)によって触媒され、ΔG° = 51 kJ.mol -1で、高い移動ポテンシャル(エノールリン酸機能)、ホスホエノールピルビン酸(PEP)を有する結合の形成をもたらします。
ピルビン酸の合成とATPの回収
リン酸基と高い移動能をもつその結合により、カップリングによる ATP 分子の合成が可能になります。この反応は Mg 2+依存性で不可逆的で、ピルビン酸キナーゼによって触媒されます。