導入
ペルオキシソーム増殖因子活性化受容体( PPAR ) は、自然に脂質に結合し、特に代謝と脂肪生成に関与する標的遺伝子の転写因子として機能する核内受容体スーパーファミリーのタンパク質です。

命名法
PPAR には、アルファ、ガンマ、デルタ (ベータ) の 3 つのタイプが確認されています。
- α (アルファ) –肝臓、腎臓、心臓、筋肉、脂肪組織などで発現します。
- γ (ガンマ) – 同じ遺伝子によって転写されますが、この PPAR は 3 つの形式で存在します。
- δ (デルタ) – いくつかの組織で発現しますが、脳、脂肪組織、皮膚で最も顕著に発現します。
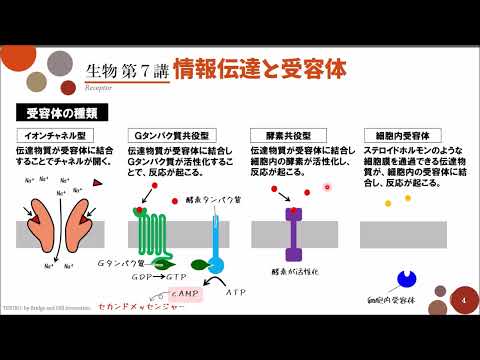
作用機序
PPAR は RXR とヘテロ二量体を形成することで、標的遺伝子の転写を活性化します。ヘテロダイマーは、PPRE (ペルオキシソーム増殖因子応答エレメントの略) と呼ばれる特定の DNA 配列に結合します。PPRE (ペルオキシソーム増殖因子応答エレメントの略) は、1ヌクレオチドの間隔で配置されたコンセンサス配列が AGGTCA である半反復部位で構成されます。
リガンド結合後の LBD の構造変化は、共調節因子の補充を誘導します。リガンドの非存在下またはアンタゴニストの存在下では、核内受容体は転写コリプレッサー ( NCoRおよび/または SMRT) と相互作用し、コアクチベーターの結合および転写複合体の開始をブロックします。コアクチベーターによるコリプレッサーの交換はタンパク質NCoEX を介して行われ、コリプレッサーのユビキチン化による抑制状態から活性状態への移行を可能にします。
活性化されたPPARは、ヌクレオソーム構造を局所的に緩和するSWI/SNFクロマチンリモデリング複合体を動員します。 2 番目のグループは、アセチル化、メチル化、リン酸化、またはユビキチン化を通じてヒストンを修飾することによってクロマチンを修飾します。 CBP/p300 は、補因子の高度に保存された LXXLL モチーフ上の LBD で PPAR 受容体に結合します。 CBP/p300 はヒストン アセチル トランスフェラーゼ (HAT)活性と転写開始因子の補充を持っています。 p160 ファミリータンパク質は、3 つのグループのコアクチベーター ( SRC1 /NCoA1、TIF2/ SRC2および pCIP/RAC3/ACTR/AIB1/TRAM1/ SRC3 ) から構成されます。これらのタンパク質は HAT 酵素活性も示しますが、Pol II 転写複合体の動員にも重要な役割を果たします。 CARM-1タンパク質とPRMT-1タンパク質も順次動員され、転写活性を増加させるヒストン メチル トランスフェラーゼです。
これらの一般的な転写補因子に加えて、PPAR には、さまざまな標的組織での作用を特異的に調節する独自の補因子があります。 PPARγ コアクチベーター 1 アルファ ( PGC1α ) は、核内受容体 PPAR、HNF4、MEF2 との相互作用を通じて、ミトコンドリア生合成を刺激し、熱産生と絶食に対する肝臓の反応 (糖新生)を制御します。 PPAR 相互作用タンパク質 (PRIP/RAP250) および PRIP 相互作用タンパク質はメチルトランスフェラーゼ活性を持っています。
構造
すべての核内受容体と同様に、PPAR は 4 つの異なる領域から構成されています。 N 末端ドメインには、リガンド非依存性トランスアクチベーター機能 (AF-1) があります。 DNA結合ドメインまたはDBDは、核内受容体スーパーファミリーに特徴的な2つのジンクフィンガーで構成されています。 PPARは、リガンド依存性トランス活性化因子機能(AF-2)を有するリガンド結合ドメインまたはLBDの前にヒンジ領域を有する。
LBD は 12 個のアルファ ヘリックスと 4 つのベータシートで構成され、大きな疎水性空洞、リガンド結合部位を構成します。リガンド-E ドメイン (LBD) アセンブリは、二量体化および転写補調節因子との相互作用のための主表面を提供します。
