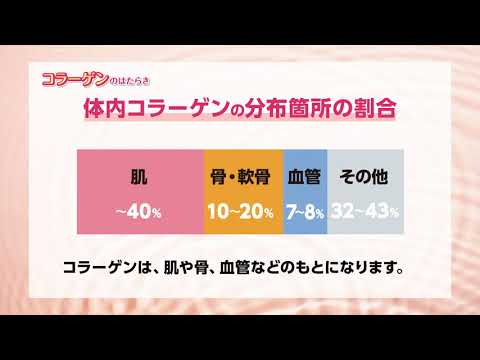
コラーゲンは繊維状の糖タンパク質であり、その役割はフレームワークに例えることができます。体内で最も豊富に存在するタンパク質です。それは結合組織細胞によって分泌されます。
結合組織にも存在するエラスチンとは異なり、コラーゲンは伸張性がなく、牽引によく耐えます。設置場所に応じてさまざまなタイプで構成されています。それは治癒過程に特に不可欠です。特定の変形の後、接着剤にすることができます。
構造タンパク質
今日の生化学者は 100,000 種類以上のタンパク質を知っています。これらの分子は多くのプロセスに関与しています。免疫学の中心となるものもあります。細胞機構が適切に機能するために不可欠な酵素活性を有するものもあります。さらに他のものは、細胞が周囲の環境と相互作用する膜受容体を形成します。一部は最終的には生物の構造に関与します。これは、動物の結合組織に必須のタンパク質であるコラーゲンの場合に当てはまります。
非常に豊富な分子
タンパク質は 2 つのカテゴリに分類されます。機能性タンパク質は生化学プロセス (酵素学、免疫学、膜受容体など) に関与しています。構造タンパク質は、その構造に寄与することで生物を形作ります。その中でも特に代表的なのがコラーゲンです。この繊維状タンパク質はすべての哺乳類の中で最も豊富で、哺乳類の体重の 25% を占めます。
コラーゲンにはさまざまな種類があります
コラーゲンは、 3 つの結合したポリペプチド鎖から構成されるタンパク質です。これら 3 つの鎖はさまざまな方法で組み合わせることができます。コラーゲンについてではなく、厳密にコラーゲンについて説明する必要があります。各タイプのコラーゲンは独自の構造を持ち、特定の臓器に存在します。たとえば、I 型コラーゲンは皮膚、腱、骨、角膜の形成に関与しており、III 型コラーゲンは心血管系に見られます。
| 親切 | 説明 | 遺伝子 |
| 私 | 脊椎動物のコラーゲンの90%。それは骨の枠組みを構成し(鉄筋コンクリートの補強材と比較して)、より一般的には通常の結合組織を構成します。骨、皮膚、腱、角膜、内臓に存在します。 | COL1A1、COL1A2 |
| Ⅱ | 椎体の髄核および眼の硝子体中の硝子軟骨(IX、X、および XI コラーゲンを含む)の基底物質に微細な原線維を形成します。 | COL2A1 |
| Ⅲ | これは、造血組織にかなりの割合で見られるレチクリン線維の構成成分です。コラーゲン状態では、特に骨格筋や血管壁に見られます。繊維は好銀性であり、陽性ではありません。 | COL3A1 |
| Ⅳ | 基底層に特有のネットワークを構成します。電子顕微鏡では縞模様はありません | COL4A1、COL4A2、COL4A3、COL4A4、COL4A5、COL4A6 |
| V | 結合組織に存在し、I型に関連します。 | COL5A1、COL5A2、COL5A3 |
| VI | 結合組織に位置し、I型に関連します。 | COL6A1、COL6A2、COL6A3 |
| Ⅶ | 基底層の構成成分。それは原線維を形成し、基底層が下にある結合組織に付着できるようにします。 | COL7A1 |
| VIII | 内皮細胞に局在します。 | COL8A1、COL8A2 |
| IX | 軟骨に位置し、II型コラーゲンと関連しています | COL9A1、COL9A2、COL9A3 |
| × | 肥大化して石灰化した軟骨に存在します。 | COL10A1 |
| XI | 軟骨に位置します。 | COL11A1、COL11A2 |
| XII | タイプ I および III と相互作用します。 | COL12A1 |
| XIII | – | COL13A1 |
| XIV | – | COL14A1 |
| XV | 散らばっている;筋肉の基底層付近。 | COL15A1 |
| XVI | – | COL16A1 |
| XVII | BPAG2とも呼ばれます。これは、特に表皮のヘミデスモソームの基底層に付着する膜貫通タンパク質です。 XVII型コラーゲン線維はα6β4インテグリンと平行に配置されています。したがって、これら 2 つのタンパク質は、基底層の固定を強化するのに役立ちます。 | COL17A1 |
| XVIII | – | COL18A1 |
| XIX | – | COL19A1 |
| XX | – | COL20A1 |
| XXI | – | COL21A1 |
| XXII | – | COL22A1 |
| XXIII | – | COL23A1 |
| XXIV | – | COL24A1 |
| XXIV | – | COL25A1 |
| XXVII | – | COL27A1 |
| XXVIII | – | COL28A1 |
天然接着剤
コラーゲンが部分的に加水分解されると、トロポコラーゲンの 3 本の鎖が解離します。このようにして、食品業界で広く使用されている製品であるゼラチンが形成されます。ゼラチンは製薬産業、化粧品、写真撮影にも使用されています。コラーゲンとゼラチンは、すべてのアミノ酸が適切な割合で含まれていないため、低品質のタンパク質とみなされます。
コラーゲンという用語は「接着剤生成者」を意味します(名前は実際には接着剤を意味するギリシャ語のkollaに由来しています)。コラーゲンの特性は4000年前にエジプト人によって利用されていました。ネイティブアメリカンは1500年前にそれを使用していました。知られている最古の接着剤はコラーゲンから作られており、その起源は 8,000 年前に遡ります。現在、美容製品や鶏の添加物として使用されています。
トロポコラーゲン
トロポコラーゲンはコラーゲンの基本単位であり、直径280nm の非対称分子に相当します。これは、3 α ポリペプチド鎖の螺旋コイル形成によって形成され、炭水化物(グルコース、ガラクトース) を運ぶ糖タンパク質です。アミノ酸は、グリシン 3 分の 1、プロリン 3 分の 1、ヒドロキシプロリン、アラニンで構成されます。 α鎖にはいくつかの分子タイプがあります。これらの鎖は 3 つのアミノ酸の反復配列で構成されており、その最初のアミノ酸であるグリシンは分子全体で反復されています。炭水化物はヒドロキシリシンに結合します。 3 α 鎖の巻きつきによりトロポコラーゲン分子が構成され、その凝集はグリシンとプロリンの間の水素結合によって確保されます。
特定のアミノ酸


トロポコラーゲン分子の組成を分析すると、多数の特異点が明らかになります。生化学者らは、残基の 3 分の 1 がグリシン残基であり、これは構造タンパク質の特徴であることを示しました。もう 1 つの特異な事実は、残基の 10% 以上がプロリンであることです。最後に、2 つの珍しいアミノ酸、 4-ヒドロキシプロリン(Hyp) と5-ヒドロキシリジン(Hyl) の存在を観察します。
特定のシーケンス
生化学者は、トロポコラーゲンを構成するアミノ酸配列にも異常があることにも注目しています。言い換えれば、そのアミノ酸配列は通常見られるものとは一致しません。実際、グリシンが 3 残基ごとに規則的に見つかることが観察されています。したがって、コラーゲンのアミノ酸配列の構造は (Gly – X n – Y n ) n となります。さらに、配列には Gly – Pro – Hyp のトリプレットが含まれることがよくあります。これらの特性はすべて、ほとんどのタンパク質には見られないという点で独特です。
水酸化とグリコシル化
水酸化
2 つのヒドロキシル化アミノ酸、ヒドロキシプロリン (Hyp) とヒドロキシリシン (Hyl) は当然のことながら、「ヒドロキシル化はペプチド合成の前に起こるのか、それとも後に起こるのか?」という疑問を引き起こします。これに答えるために、生化学者はこれらのアミノ酸のいずれかをコードするアミノアシル tRNA を探しました。何も見つからなかったため、彼らは特定のプロリンおよびリジン残基の水酸化は翻訳後のプロセスであると結論付けました。実際、2 つの酵素がこの反応を触媒します。これらはそれぞれプロリルヒドロキシラーゼとリシルヒドロキシラーゼです。どちらもアスコルビン酸塩(またはビタミンC)の存在下でのみ作用でき、活性中心に含まれる第一鉄(Fe 2+ )が第二鉄(Fe 3+ )の形で不活化されるのを防ぎます。重度のアスコルビン酸欠乏は、コラーゲン生成の欠陥によって引き起こされる病気である壊血病を引き起こし、その結果、組織が弱体化します。船員は、長い航海中に果物や野菜を食べることができず、しばしば壊血病の犠牲者になりました。
ヒドロキシル化可能なプロリン残基は必ずグリシン残基の左側に位置します。他の残基はヒドロキシル化できません。リジン残基についても同様です。
(アミノ末端) NH 3+ – … – Pro – Gly – … – COO – (カルボキシ末端)
(アミノ末端) NH 3+ – … – Lys – Gly – … – COO – (カルボキシ末端)
ごくまれに、特定のプロリン残基が炭素3 で水酸化され、炭素 4 では水酸化されないことがあります。この水酸化に関与する酵素はプロリルヒドロキシラーゼではなく、水酸化は先ほど定義した規則に準拠しません。
グリコシル化

生化学者たちは、トロポコラーゲンの構造を研究することによって、一部のヒドロキシリシン残基が糖残基(2-グルコシル-ガラクトース)を持っていることも示しました。コラーゲンが短い炭水化物に共有結合しているという事実により、それは糖タンパク質になります(プロテオグリカンとは対照的に、小さなペプチドに結合した炭水化物です)。ガラクトースとヒドロキシリシン残基のヒドロキシル基との間の共有結合の形成は、ガラクトシルトランスフェラーゼによって触媒されます。グルコースのそれはグルコシルトランスフェラーゼを使用します。
二次構造
ヘリカルシリンダー
トロポコラーゲンの空間構造は、長さ 3000 Å、直径 15 Å の 3 本の縞模様のらせん状円柱の構造です。したがって、このタンパク質は、機能性タンパク質の古典的な球形をしていません。細長い茎です。円柱の各バンドは、それ自体がらせん状に折り畳まれたポリペプチド鎖です。 3 つのチェーンすべてが特に強力な組紐を形成します。実際、直径1ミリのコラーゲン繊維を切断するには10kgの荷重がかかります。
左らせん構造
各ポリペプチド鎖はらせん状に折りたたまれていると言いました。この二次構造はαヘリックスとは無関係です。これも二次構造の一種です。これは、多数のプロリン残基のピロリドン環の反発によって曲がりくねったらせんです。ピロリドン環は、強い立体障害により互いに反発します。得られたヘリックスは、残基あたり 2.9 Å の距離を測定できたため、α ヘリックス (残基あたり 1.5 Å) よりも充填度が低くなります。最後に、αヘリックスとは異なり、鎖内の水素橋は観察されないことに注意する必要があります。それにもかかわらず、私たちはカテナリー間の水素橋を発見しました。水素結合には、グリシン残基 (プロトン供与体) とカルボキシル基 (プロトン受容体) が関与します。鎖が交差する場合には立体障害を軽減する必要があります。これは、最も障害の少ないアミノ酸であるグリシン残基の存在によって可能になります。他の側鎖は三重らせん構造の外側に放り出されます。
協力的な相互作用
トロポコラーゲンの溶液を加熱すると、T m (アングロサクソンの用語で融解温度) と呼ばれる、特定の温度でらせん構造の突然の崩壊が観察されます。この実験は、水素架橋によって三重らせん構造が確実に維持されることを示しています。これらの弱い(つまり、非共有結合的な)相互作用は、分子の挙動を大きく変えるのに十分な数です。私たちは驚くべきではありません。クモによって生産される絹の構造タンパク質であるフィブロインは、その弾力性と耐性を、協力的な相互作用を構成する多数の水素橋の存在に負っています。同様に、水の独特の性質、特にその異常に高い沸点は、多数の水素橋の存在の直接の結果です。融解温度 T mの測定は、溶液の粘度の変化と回転光学分散(DOR) の変化を示すという事実によって可能になります。
水酸化の重要性
コラーゲンで観察されるヒドロキシル化により、安定性が向上します。したがって、その数は組織の種類と体温によって異なります。一部の組織は他の組織よりも高い水酸化率を持っています。これは特に胎盤と皮膚に当てはまります。
| プロとハイプ (∴) | 融解温度(℃) | 体温(℃) | |
| カーフスキン | 232 | 39 | 37 |
| サメ肌 | 191 | 29 | 24~28 |
| カビヨ | 155 | 16 | 10~14 |
コラーゲン生合成

プロコラーゲン
コラーゲンは細胞自体には存在しません。その合成が細胞内に存在する前駆体であるプロコラーゲンから行われるのはこのためです。これは、アミノ末端側が 15 kDa、カルボキシ末端側が 30 kDa の追加のポリペプチド構造によって延長された、はるかに長い鎖で構成されています。これらの構造はありふれたものであり、プロペプチドと呼ばれます。カルボキシ末端には、安定化の役割を持つ鎖間ジスルフィド架橋があります。架橋はアミノ末端の架線内にあります。
プロコラーゲン
プロコラーゲンは、直線らせん状の 3 つのアルファ コラーゲン タンパク質の集合体です。プロコラーゲン線維は、コラーゲン原線維と呼ばれる構造を形成します。いくつかのコラーゲン原線維が集合してコラーゲン線維を形成します。原線維は、プロコラーゲン線維の間隔とオフセットにより、暗相と明相が交互に現れます。
小胞体では
3 本のプロコラーゲン鎖が合成され、小胞体の内腔に移動します (これは転座と呼ばれます)。三重らせんを含む領域には、シグナルペプチドと 2 つのアミノ末端およびカルボキシ末端のプロペプチドが隣接しています。これらはタンパク質の成熟を助けます。
翻訳後修飾
小胞体に入ると、シグナルペプチドは切断されます。次に、いくつかのプロリンとリジンの水酸化が起こります。カルボキシ末端プロペプチドは N-グリコシル化によって修飾されますが、一部のヒドロキシリシンは O-グリコシル化されます。さまざまな翻訳後修飾と 3 つの鎖のカルボキシ末端プロペプチド間のジスルフィド架橋の形成により、それらの整列が可能になります。その後、三重らせんはアミノ末端に向かってジッパーのようにとぐろを巻きます。
ゴルジ体では
最後に、ゴルジ体では、三重らせんの両側に非らせん領域があり、プロペプチドはエキソサイトーシスによって細胞外環境に放出されます。次に、酵素であるプロコラーゲン ペプチダーゼが2 つのプロペプチドを切断し、成熟したトロポコラーゲンの形成を可能にします。最後に、トロポコラーゲンの束が細胞表面近くに集まり、相互接続して成熟したコラーゲン線維を形成します。
コラーゲン線維の形成
成熟したコラーゲン線維を形成するには、トロポコラーゲンの束が集まる必要があります。さまざまな生化学的プロセスがこれにつながります。たとえば、酵素であるリシルオキシダーゼは、トロポコラーゲンのいくつかの束への自発的架橋の起点となるリジン残基のアミン基をカルボニル基に置換することを可能にします。したがって、アルドール縮合によって分子の架橋を実行します。

別のメカニズムには、ヒドロキシピリジニウム(またはヒドロキシピリジノリン) 架橋を介して 3 つのポリペプチド領域を接続することが含まれます。その後、680 Å の周期性を持つはしご構造が形成され、電子顕微鏡で観察できます。異なるトロポコラーゲン分子間の空いた領域は、式 Ca 10 (PO 4 ) 6 (OH) 2のヒドロキシリン酸カルシウム、ヒドロキシアパタイトを使用して充填されます。トロポコラーゲンとヒドロキシアパタイトの規則的な連続は、電子顕微鏡で観察できる縞模様の起源にあります。
コラーゲンの分解
コラゲナーゼ
コラーゲンの分解は難しく、特定の酵素であるコラゲナーゼ(これらはメタロプロテアーゼです)を必要とします。例えば、細菌由来のコラゲナーゼ(例えば、種名が「組織破壊者」を意味するClostridium histoliticum )は、各鎖を 200 以上の点(そのうち X-Gly-Pro-Y )に切断することができます。両生類などの他の生物は、単一の切断を行うことができる非常に特異的なコラゲナーゼを持っています。
コラーゲン関連疾患
すでに説明した壊血病に加えて、脆性骨疾患を含む多くの病気がコラーゲン合成の欠陥に関連しています。遺伝性膠原病の記事を参照してください。