導入

ヒト胚形成(または胚発生)とは、受精卵の段階から出生までのヒト胚の発生過程を指します。
胚発生の段階
受精
接合子または卵子は、女性の配偶子(卵母細胞) と男性の配偶子 (精子) の結合から生まれます。配偶子は生殖細胞の減数分裂によって生成されます。したがって、雄の配偶子は一倍体です。雄の配偶子は、それを生成した生殖細胞の相同染色体の各対の 2 つの染色体のうち 1 つだけを含みます。女性の配偶子には相同染色体が 1 つだけ含まれています (受精があった場合にのみ減数分裂が完了し、その後二倍体になります。卵母細胞は卵子になります)。
精子は、卵母細胞の周囲の透明帯内の糖タンパク質に結合します。まず ZP3 に結合し、先体反応が引き起こされます。精子の原形質膜は、前方部分から始まり、論理的に前後の勾配に沿って先体の外膜と融合し、3 つの層が存在する赤道帯を残します。 (先体の外膜と内膜、および原形質膜) が残ります。先体嚢が外側に開くと、酵素の放出が生じます。その中には、アクロシンとヒアルロニダーゼが挙げられます。これら 2 つの酵素は、糖タンパク質 ZP1 を ZP2 および ZP3 から分離することを可能にし、精子が卵母細胞に進むために不可欠です。透明帯内のすべての精子は先体反応を行っています。 ZP2 は精子が結合する 2 番目の糖タンパク質であり、精子の移動も促進します。
精子は卵母細胞の微絨毛ゾーンに近づき(微絨毛ゾーンは、過剰な遺伝物質を含む第2極体の排出のために確保されています)、2つの細胞の融合によってカルシウム波が引き起こされ、その結果皮質顆粒が押し出されます。 、リソソームが負荷され、ZP2糖タンパク質を精子に結合できないZP2Fに変換します。このメカニズムにより、多精子のブロック、つまり単一の精子が卵子に受精することが保証されます。
一倍体配偶子の融合には、雄と雌の 2 つの前核 (一倍体核) の結合が含まれます。これら 2 つの前核の接近と融合の段階は、それぞれ合婚と両性混合に対応します。 「卵子」という用語は、出会いの過程で 2 つの前核を含む場合にのみ、「卵母細胞」という用語の代わりに使用できます。
雄と雌の相同染色分体は接合子内で結合します。したがって、それとその子孫には「混合」染色体が含まれています。これは、個人の多様性と独自性を確保するのに役立ちます。さらに、これらの遺伝子は、次世代の最適な遺伝子混合を確保するために、ある染色分体から別の染色分体へ受け継がれることによって交差 (いわゆる交叉) することができます。それ以来、受精卵には、細胞の分化と分化という複雑なプロセスを経て、生物に変化するために必要なすべての情報が含まれています。
分割: 卵から桑実胚まで
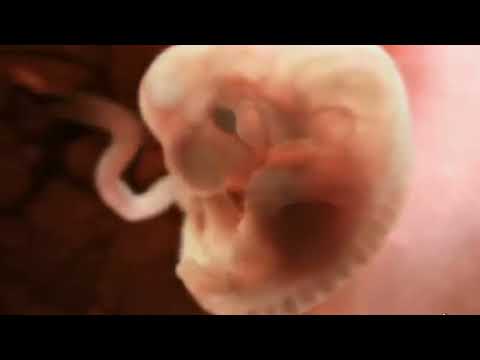
受精卵は、卵管と子宮に移動する間に一連の細胞分裂を行います。このプロセスはセグメント化、または切断と呼ばれます。このセグメンテーションは完全、非同期、非対称です (生成された 2 つの娘細胞は同じ量の細胞質物質を保持していない可能性があり、サイズに顕著な違いが生じます)。受精卵は、いくつかの連続する古典的な有糸分裂によって、最初に2つの娘細胞に、次に3つに(最大の割球が最初に分裂する)、次に6と8というように分割され、すぐに桑実胚(桑実胚のように見える)という名前を持つ細胞塊に到達します。小さなブラックベリー、その名前が示すように)。 2 番目の「卵割」という用語は、最初の連続する分裂中に受精卵のサイズや体積が増加しないため、完全に刺激的です。 4~8細胞期までの最初の細胞分裂は、娘細胞間に顕著な形態学的差異を伴わない。
一般に受け入れられている年表は次のとおりです。発生初日に 2 個の割球が受精後すぐに出現し、発生 2 日目に 4 個の割球、発生 3 日目に 8 個の割球、4 日目に 16 個の割球 (桑実胚) が出現します。 。 5日目には、通常、胚は子宮の上部に到達し、そこで並置されて着床します(着床または卵着床の現象)。
8 細胞期から 16 細胞期にかけて、圧縮現象により桑実胚内に新しい細胞分布が生成され、胚分化の最初のイベントが開始されます。
受精後 4 日目の終わりに、空洞形成現象により桑実胚が空洞になり、液体内容物を含む空洞が現れます。これが将来の胚盤腔です。
入れ子
子宮内膜への卵子の着床は、胚芽の反対側にある細胞栄養芽層(現在孵化した透明帯の内部部分)の細胞によって行われます。卵子は、発育の5日目から7日目(無月経の19日目から21日目)の着床期間中に子宮底に着床します。
栄養膜細胞は、細胞クラスターを球の一部に押し込む液体を分泌します。このようにして合体栄養膜細胞が分泌され、リソソームが蓄積され、そのメタロプロテイナーゼ機能のおかげで子宮内膜にどんどん広がります。
絨毛膜は卵を取り囲む複合構造であり、内部から周縁まで、胚外間葉、細胞栄養膜、そして最後に上記の合胞体栄養膜の 3 つの層で構成されています。キャビテーションにより、母親の血液室が出現し、12日目から母親から酸素を受け取ることができるようになります。血液循環( 21日目までは組織栄養性) は絨毛膜絨毛を介して起こり、 14日目までは主要な状態を保ちます。絨毛膜は子宮に胎児を迎えるように指示するホルモンを分泌します。特定の物質は母体の免疫応答を制御し、(移植された臓器の場合のように)胚の拒絶反応が起こらないようにします。
発芽
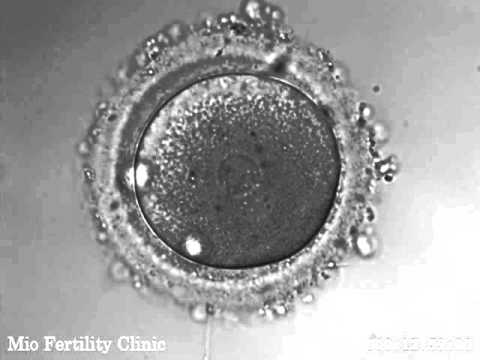
人類では、接合子 (または卵子) はアレシス (またはメタレシス:胎盤が卵黄の代わりとなる) タイプです。卵黄は少量ではありますが、細胞質全体に均一に分散しています (詳細については、接合子に関する記事を参照してください)。 )。
発芽の最初の段階で、受精卵は有糸分裂によって割球に分割されます。すべてのアレシサスの接合体と同様に、このプロセスは完全芽球的で放射状です。最初の分裂は、接合子の極を通過する南面に沿って起こります。次に、結果として生じる 2 つの割球のうちの 1 つも南面に沿って分裂します。一方、もう一方の割球は赤道面に沿って分裂します。連続的な分裂が進むにつれて、凝集したままの細胞が桑実胚を形成します。この外側の細胞は、胎盤の表層を構成する栄養膜を形成します。内部細胞は桑実胚の一方の極に集合し、内部細胞塊および空洞である胚盤尾を形成します。
この段階では、桑実胚は胞胚または胚盤胞になります。
原腸形成
導入
原腸形成のプロセスは、胚を生成し、その発達をサポートする細胞構造を確立します。
この目的を達成するために、胚盤胞細胞は分化、移動、再配置されて以下を形成します。
- 胚組織(胚層)。
- 胚外組織(支持構造)。
人類の原腸形成は「移民」タイプです。胚盤は胚ボタンのレベルで形成されます。個々の細胞は、外胚葉、内胚葉、中胚葉への分化と同時に椎間板を横切って移動します。このプロセスは、すべての哺乳類だけでなく、その祖先である爬虫類や鳥類にも共通しています。
胚と栄養機能
胚盤胞の頂端では、胚芽細胞または胚ボタンと呼ばれる内部細胞のクラスターが形成されます。ここから胚の発生が始まります。
胚盤胞の末梢細胞の栄養機能がより明確になります。それらは栄養膜を構成し、外側の合胞体栄養膜 (多細胞細胞質) と内側の栄養膜細胞に分かれます。
胚芽細胞は、内芽細胞 (下芽細胞) と外芽細胞 (胚盤葉上層) に分かれます。これと栄養細胞膜との間で羊膜腔が空洞になります。胚盤葉上層と胚盤葉下層は一緒になって胚盤を形成します(胚盤葉下層は最終的な葉を生成しないため、胚葉盤について話すことはできません)。
胚盤葉上細胞は羊芽細胞に分化し、羊腔の内壁を覆います。
合胞体栄養膜では、空胞が開き、互いに接続して毛細管を形成し、毛細管が子宮組織内に伸びて母体洞を構成します。液胞は血液で満たされる空隙を形成します。これらの血管は子宮胎盤循環の基礎です。
最初に細胞が栄養膜層の内壁に沿って下芽細胞から遊走し、胚盤腔を一次卵黄嚢に変換する薄い膜(胚体外内胚葉、「ホイザー膜」とも呼ばれる)を形成します(D-11)。
中胚葉の出現と羊膜の成長
胚体外小胞体(非常に緩い無細胞組織で、「胚体外間葉」とも呼ばれる)は、ホイザー膜と栄養細胞層の間に現れます。そこで液胞が開き、融合して液体で満たされる胚体外腔を形成します。小胞体は膜によって生成されると考えられています。
中胚葉細胞は、胚盤尾の尾側部分から生じる分化した胚盤葉上細胞であり、小胞体を通って移動し、付着することによって胚外腔の壁を覆います。この胚外中胚葉は、細胞栄養芽層、羊膜腔、および胚茎のレベルでは体葉胸膜と呼ばれ、一次および二次卵黄嚢の胚体外内胚葉のレベルでは内臓胸膜と呼ばれます。
胚外内胚葉細胞の新たな移動は、一次卵黄嚢の壁に沿って移動し、次に内側に移動することによって、以前の細胞を押し戻します。一次卵黄嚢は、胚外腔の圧力で収縮し、一次卵黄嚢と、急速に退縮する残存物である体腔外嚢胞に分裂します。新しく形成された 2 つの小胞は、内臓胸膜によって外側が覆われています。
羊膜腔の成長の結果、胚外腔は縮小します。体腔液が失われ、羊膜と胚体外腔の胚体外体胸膜が融合します。
三皮胚
胚盤葉上細胞は胚盤の背側縦軸に向かって集まり、「原始線」と呼ばれる縁を形成します。これにより、胚盤葉上細胞が椎間板の中に沈んで分散し、縁が溝に変わります。胚盤葉上細胞には三重の運命があります。
- 一部は胚盤葉下細胞の間に挿入され、胚体内胚葉を形成します。
- 他のものは胚盤葉上層と胚盤葉下層の間で停止し、最終的な胚性中胚葉に分化します。
- 胚盤葉上層に残ったものは、最終的な胚外胚葉になります。
この段階では、胚は三皮体になります。これには外胚葉、内胚葉、中胚葉が含まれており、これら 3 つはすべて胚盤葉上層から形成されます。
胚盤の構造
原始線の外観は、椎間板の左右対称性だけでなく、その軸の尾頭側の向きも定義します。 「ヘンセン ノード」は軸の中央付近に位置するくぼみで、プリミティブ ラインが終了します。
胚盤の尾側端では、外胚葉の領域が内皮にくっついたままになっています。これは総排出腔膜であり、陥入した原腸胚の原孔の相同体です。同様に、咽頭(または口腔咽頭)と呼ばれる膜が頭蓋端に向かって形成されます。その破裂により、原始的な口が咽頭と接触し、羊水が流れ込む移行管、つまり口腔が残ります。
神経形成
神経形成プロセスの開始時に、コードが確立されます。これは、ヘンセン結節レベルでの中胚葉細胞の増殖から始まります。次に、細胞は頭蓋極に向かって移動し、外胚葉と内胚葉の間の円板の軸に中空の管、脊索突起 (または管) を形成します。最初に、脊索管は内皮と融合し、こうして脊索板を形成します。その後、内皮から分離して固体のコイル、つまり弦そのものになります。このプロセス中に、羊膜腔と二次卵黄嚢の間の唯一の(一時的な)連絡である神経腸管が数時間( 18日目と19日目の間)存在することにも注目します。弦の形成に必要なこのチャネルの形成は、上記以外の場合にも、重篤な、しばしば致死的な病状を引き起こします。
弦の存在により、上にある外胚葉からの神経板の形成が誘導されます。これは頭蓋極で幅が発達し、そこに脳が形成されます。尾極に向かって、それは側溝の形をしており、閉じるとき、その端は脊髄の起点にあります。
要約すれば:
- 前脳、中脳、後脳の形成。
- 神経溝の完全な腸重積と神経管の分離。
- 椎間板の極に向かって神経管が伸びる。
- 神経堤の出現。その遊走細胞が体の多数の組織を形成します。
メタメライゼーション
その間に、胚内の中胚葉が分化します。脊索の両側に位置する傍軸中胚葉は圧縮され、頭蓋極から始まる 44 対の体節を形成します。個別化プロセス (メタマー化) の間、これらは中間層によって中胚葉に接続されたままになります。側板は、外胚葉を覆う体胸層と内胚葉を覆う内臓胸層の 2 つの層に分かれています。
体節からは次のことがわかります。
動静脈系が確立され、胎児と母親の間で以下のような最初の交換が可能になります。
- 栄養膜絨毛レベルでの胚体外循環、
- 原始的な胚内循環。
胚の境界
それは胚とその付属物の分離です。胎児の輪郭と形状は、羊膜と卵黄嚢の間の成長の差異によって引き起こされます。これは事実上もはや発達せず、羊膜が四方八方から溢れ出ます。
側方では、羊膜は胎児の椎間板、つまり神経管、索、胚内の中胚葉、そして原始腸となる卵黄の一部を覆っています。さまざまな層の端が椎間板の基部で出会い(臍帯を形成します)、それらは融合して胚の本体を閉じ、三次元になります。しかし、腸と卵黄嚢の間には、卵黄管というチャネルが一時的に残ります。縦方向に、胚は凸状の形状をとります。膜は約180°回転します。心原性中芽細胞は、最初は咽頭膜の前にあり、腹側の位置に移動します。
全体的な構造が整うと、さまざまな器官が発達します。
残りは形態形成の記事で開発されます。